
Перейдем теперь к описанию предполагаемых механизмов первичного поглощения пигментами энергии света в преобразовании ее в энергию химических связей.
Световые потоки обладают двоякой природой. По некоторым признакам они обладают свойствами потока электромагнитных волн, длина которых в видимой части изменяется от 380 до 720 мμ (в этой же части спектра сосредоточены лучи, активные при фотосинтезе). С другой стороны, по некоторым признакам световой поток может быть охарактеризован как поток определенных порций энергии, называемых фотонами или квантами энергии. Энергия квантов света с более короткой длиной волны больше, чем энергия квантов света с более длинной волной.
Молекулы веществ, подвергаемых превращению, в том числе и пигментов фотосинтетического аппарата, поглощают свет квантами. Каждая молекула в исходном ее состоянии может поглотить только один квант. При этом меняется энергетический уровень состояния молекул и, в частности, если говорить о молекуле хлорофилла, то один из ее электронов возбуждается и переходит на более высокий энергетический уровень.
В разных условиях судьба этого электрона может быть различной. Этот электрон может потерять приобретенную им энергию, переходя с одного более высокого энергетического уровня к более низкому. При этом электрон может отдавать энергию в виде кванта света (флуоресценция) или тепла. В этом случае полезного запасания энергии при поглощении кванта света не произойдет. Такое явление наблюдается обычно в простых растворах хлорофилла, который при освещении флуоресцируют ярко-красным светом или нагревается.
Другое дело, в структурированном фотосинтетическом аппарате. Здесь электрон из молекулы хлорофилла, поднятый на высокий, уровень теми или иными, пока еще не вполне ясными, путями, переходит, сохраняя приобретенную энергию, к активным центрам, где он может приниматься активными катализаторами, восстанавливая их и обогащая энергией.
Работами последних лет установлено, что в процессе фотосинтеза осуществляется, по крайней мере, два фотохимических акта, происходящих при участии молекул двух форм пигментов, возможно хлорофиллов а и б.
Ближайшим результатом превращения обогащенного энергией выбитого из молекулы хлорофилла а электрона будет захватывание его таким активным веществом, как фередоксин (железосодержащий белок). Последний в свою очередь передает электрон со значительным запасом энергии тем переносчикам водорода, о которых мы говорили выше и которые носят название пиридиннуклеотидов. Принимая электрон, а вслед за ним положительно заряженный ион водорода, пиридиннуклеотиды приобретают свойства чрезвычайно активного восстановителя и используются в дальнейшем как восстановители в цикле превращения углерода, о котором мы уже сообщали выше. Другие электроны, выбиваемые таким же образом из молекул хлорофилла а, также переходят по нисходящей энергетической лестнице от одного переносчика к другому, обогащая их энергией и, в конце концов, отдавая ее на образование богатых энергией молекул аденозинтрифосфорной кислоты.
Так, по образному выражению американского исследователя Арнона, энергия кванта света, переносимая возбужденным электроном, выбитым из молекулы хлорофилла, разменивается на ходовую валюту в виде восстановленных пиридиннуклеотидов и аденозинтрифосфорной кислоты. Эти обычные для биологических реакций переносчики водорода и энергии используются и в процессе фотосинтеза как агенты для восстановления окисленного углерода, карбоксильных групп, нитратов, сульфатов, для активации аминокислот и простейших углеводов, в результате чего образуются сложные углеводы, белки и т. д.
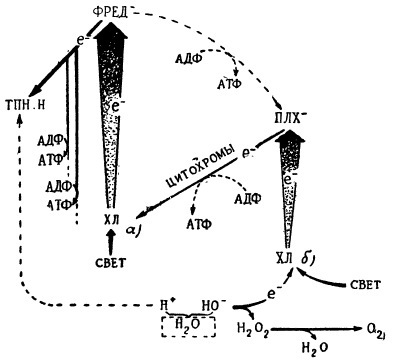
Схематическое изображение переноса электрона и водорода от воды на пиридиннуклеотиды, сопровождающегося запасанием энергии (ФРЕД — фередоксин, ПЛХ — пластохинон. По Арнону)
Что же происходит с молекулами хлорофилла а, которые, поглотив кванты света, потеряли электроны. В этом случае в активных слоях хлоропластов, где находятся молекулы пигментов, образуются электронные вакансии — «дырки». Вакансии оказываются способными активно поглощать электроны из какого-либо другого источника. При этом молекулы хлорофилла восстанавливают свою активность и способность опять поглощать квант света и направлять его энергию и электроны в фотосинтетический цикл.
Такой источник пополнения утраченных хлорофиллом и электронов — вода. По последним представлениям этот дополнительный поток электронов обусловлен работой другой формы хлорофилла — хлорофилла б. В этом случае электрон из молекулы хлорофилла б при поглощении кванта света также переходит на высокий энергетический уровень и через несколько ступеней с помощью окислительных ферментов передается на хлорофилл а, возмещая отданные им ранее электроны и восстанавливая опять работоспособность пигмента. Что касается молекул хлорофилла б, то он принимает электрон от воды через гидроксильный ион и тем самым восстанавливает свою активность. Потеряв электрон, гидроксильные группы образуют перекись, которая разлагается с выделением свободного кислорода. Возможно, что участниками этого переноса кислорода до выделения его в виде молекул газа (O2) выступают каротиноиды.
Так по современным представлениям при участии двух форм хлорофилла и двух фотохимических реакций осуществляется «сквозной поток» электронов, движимый энергией света от воды через молекулы двух форм хлорофилла на образование восстановительной силы фотосинтеза в виде пиридиннуклеотидов и АТФ с устойчивым запасанием в них энергии. Эта энергия затем передается на восстановление углерода, нитратов и на образование прямых продуктов фотосинтеза: углеводов, белков и т. д.
Осуществление всего цикла фотосинтеза предполагает восстановление углерода, активизацию воды и в ряде случаев активирование и восстановление сульфатов, а также нитратов. Для осуществления цикла необходимо не менее 3—4 молекул АТФ и четырех молекул восстановленных пиридиннуклеотидов в расчете на одну молекулу усвоенной углекислоты. Чтобы они образовались в описанном выше процессе мобилизации электрона и водорода, должно произойти 7—8 элементарных фотохимических актов поглощения соответствующего числа квантов энергии света. Это значит, что в наилучших условиях фотосинтез может идти примерно с 8-квантозым расходом, т. е. с поглощением и полезным использованием на фотосинтетическую работу 8 квантов света для восстановления одной молекулы углекислоты.
Однако это возможно лишь тогда, когда вся система работает безукоризненно слаженно, когда все реакции точно согласованы между собой и производительность активных агентов каждой ступени фотосинтеза не лимитирует производительной способности агентов других звеньев. При этом все звенья системы тесно взаимосвязаны и работа их взаимно обусловлена. Доказать это несложно. Хорошо освещаемый и снабжаемый углекислым, газам лист может осуществлять интенсивный фотосинтез, т. е., интенсивно поглощать углекислый газ и выделять кислород. Но стоит быстро переместить лист в атмосферу, лишенную углекислоты, и моментально прекратится выделение кислорода, хотя в листе есть и вода, из которой может выделяться кислород, и хлорофилл и подается энергия света. Однако выключение из системы конечного потребителя электронов и водорода — CO2 почти моментально нарушает действие всей цепи в целом.
Мы привели пример лишь крайнего воздействия на работу фотосинтетического аппарата, хотя он представляет собой сложный процесс, проходящий несколько стадий. Естественно предполагать, что уклонение от нормы в работе любого звена этой сложной системы или отставание одного звена от работы других звеньев может и должно определенно сказываться как на количественных, так и на качественных результатах работы фотосинтетического аппарата. Вместе с тем нормальная работа аппарата обусловлена не только наличием или отсутствием тех или иных активных элементов, не только изменением количественных отношений их содержания, но и особенностями структурной их организации. А она также чрезвычайно изменчива.
Кроме того, надо иметь в виду, что хлоропласты неотъемлемая часть фотосинтезирующих клеток. Они находятся в тесном взаимодействии со всеми ее частями. Состояние же и деятельность этих частей определяется состоянием и деятельностью растений в целом и, в частности, условиями, которые так или иначе благоприятствуют или не благоприятствуют его росту.
Если растение хорошо обеспечено питанием, энергично растет, оно — активный потребитель продуктов, создаваемых фотосинтетическим аппаратом, и тем самым активизирует его деятельность. Если же процессы роста подавлены вследствие недостатка влаги, элементов минерального питания, сильных отклонений от нормального температурного режима, то потребление продуктов фотосинтеза резко снижается. Усваивающие новый продукт клетки и хлоропласты перезаполняются продуктами фотосинтеза: сахарами, крахмалом, и фотосинтез подавляется.
Читатель представляет себе, что работа фотосинтетического аппарата растений чрезвычайно сложна и зависима от многих условий. При этом положении невозможно говорить о каком-то определенном для всех условий КПД зеленого листа. Очевидно, дело должно сводиться к тому, чтобы определить максимально возможный КПД, вытекающий из самой природы механизма фотосинтеза, и в оптимальных условиях его работы, а затем устанавливать фактические КПД в разных условиях, выяснять причины отличия их от теоретически возможных и изыскивать способы их повышения.
Мы уже говорили о том, что для осуществления полного цикла фотосинтеза в максимально благоприятных условиях нужна затрата не менее четырех молекул, восстановленных пиридиннуклеотидов и 3—4 молекул АТФ Образование их может быть связано с поглощением 8 квантов света. Округленно 8-квантовый расход определяет максимально возможный КПД фотосинтеза растений. Энергетический выход фотосинтеза в этих условиях мы рассмотрим ниже.